中国科学院遗传与发育生物学研究所杨维才研究组鉴定了一个水稻花器官发育异常的突变体,用改良的Mutmap方法定位了OsPID的变异位点,并用CRISPR/Cas9基因编辑技术和遗传互补试验进行了验证。与拟南芥AtPID相比,OsPID除了分布在细胞膜上,还存在于细胞核及细胞质。该研究进一步发现OsPID与OsPIN1a、OsPIN1b及OsMADS16等具有相互作用。总结本研究及前人研究结果,论文提出了OsPID调控水稻花器官发育的模型。首先,像在拟南芥中一样,OsPID通过磷酸化PIN1而调控生长素的运输,改变生长素的极性分布,调控水稻花器官的产生和发育;其次,OsPID还通过与OsMADS16、LAX1等转录因子相互作用参与调控水稻花器官发育;另外,OsPID与LAX1/LAX2互作,通过控制水稻的分枝和分蘖而影响产量。OsPID的过量表达可使穗分枝数、穗粒数及单株产量增加,但不影响千粒重和穗数,这说明OsPID在水稻高产育种中具有潜在的应用前景。该研究结果为水稻花器官发育的分子机制提供了新的认识,为水稻高产育种的遗传改良提供了理论基础。
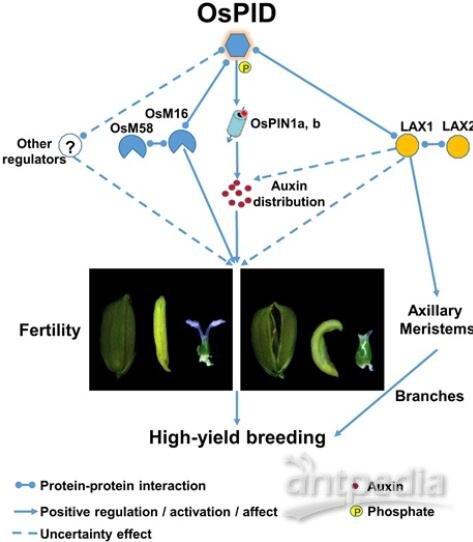
图:OsPID调控水稻花器官发育的分子机制模型
水稻是世界上一半以上人口的主粮,其产量主要受每穗粒数、每株穗数、千粒重等影响。其中每穗粒数与每穗颖花数密切相关,因此颖花的发生和发育直接影响了水稻的产量。在拟南芥中,PINOID(PID)可以通过调控生长素外流载体PIN家族蛋白的亚细胞定位来调节生长素的分布(Friml et al.,2004;Lee and Cho,2006)。AtPID是控制花原基形成的基因,其通过生长素途径参与调控双子叶的花器官发育的机理已有很多报道,而该基因在单子叶植物的生物学功能还不清楚。之前有研究发现,将OsPID在水稻中进行过量表达会引起雄蕊数目减少、雌蕊数目增加的异常表型(Morita and Kyozuka,2007)。尽管最近的两篇报道表明,OsPID功能缺失会导致水稻雌蕊和花药发育异常(He et al.,2019;Xu et al.,2019),但PID调控水稻花器官的分子机制尚不清楚。
该研究成果于1月17日在Plant Biotechnology Journal杂志上在线发表(DOI:10.1111/pbi.13340)。杨维才研究组博士后吴华茂为文章第一作者,杨维才为通讯作者。该项研究得到中科院战略性先导专项和国家自然基金委项目的经费资助。